Механизм синтеза АТФ при участии АТФ-синтазы получил название вращательный катализ (роторный катализ, ротационный катализ).
Фермент АТФ-синтаза состоит из множества белковых цепей, формирующих два больших компонента:
- компонент Fо (олигомицин-чувствительный) – его функция каналообразующая, через него выкачанные наружу ионы водорода устремляются в матрикс,
- компонент F1 (fraction 1, англ. - часть 1) – его функция каталитическая. Именно он, используя энергию протонного градиента, синтезирует АТФ.
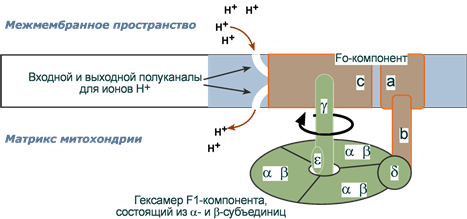
Строение компонента Fо
Данный компонент погружен в мембрану, является интегральным белком цилиндрической формы, образован субъединицами типов a и b, и 10-12 субъединицами типа с, собранными в единый комплекс. В каждой из c-субъединиц есть отрицательно заряженные центры связывания протонов – остатки аспарагиновой кислоты. Эти центры взаимодействуют с полуканалами для ионов H+, открывающимися наружу (в межмембранное пространство), и внутрь (в матрикс).
a- и b-Субъединицы являются структурными. Их задача - обеспечить прикрепление к мембране F1-компонента.
Строение АТФ-синтазы
Строение компонента F1
Молекулы этого компонента состоят из девяти субъединиц пяти различных типов (3α, 3β, γ, δ, ε). Основной функциональной субъединицей F1-компонента является гексамер, состоящий из 3α- и 3β-субъединиц. Через δ-субъединицу гексамер присоединен к b-субъединице (Fo), которая в свою очередь зацеплена в мембране за a-субъединицу Fo-компонента. Таким образом, гексамер 3αβ фиксирован и неподвижен. Каталитический центр, в котором и происходит синтез АТФ, находится в β-субъединице.
γ-Субъединица одним концом жестко связана с комплексом c-субъединиц (Fo), другим концом она входит внутрь гексамера 3αβ. С ней дополнительно связана минорная субъединица ε.
Взаимодействие F1 и Fo
Синтез АТФ происходит за счет энергии, высвобождающейся при прохождении протонов через каналы комплекса Fo.
Протоны из межмембранного пространства достигают своих центров связывания на с-субъединицах (Fo) через входной полуканал и прикрепляются к аспартату, нейтрализуя его отрицательный заряд. Лишенная заряда с-субъединица меняет свою конформацию и принуждает Fo-комплекс вращаться вокруг своей оси, доставляя протоны к другому полуканалу, который направлен уже в матрикс митохондрий. Поскольку концентрация ионов H+ в матриксе низка, то они легко отрываются от аспартата и уходят внутрь, а оставшийся, уже заряженный, аспартат не позволяет комплексу вращаться в обратном направлении.
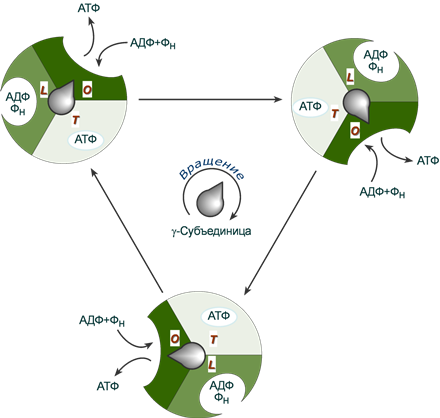
При связывании ионов H+ происходит поворот не только комплекса c-субъединиц Fo-компонента, но и жестко связанной с ним γ-субъединицы F1-компонента. Стержень γ–субъединицы проворачивается внутри неподвижного гексамера 3αβ и при каждом повороте на 120° она поочередно вступает в контакт с каталитическими β-субъединицами, что меняет их функционирование.
Взаимодействие γ-субъединицы с β-субъединицами при синтезе АТФ
β-субъединицы могут находиться в трех конформациях, выполняющих разную функцию:
- loose, L (англ. слабо связано) – удерживает АДФ и ион фосфата,
- tight, T (англ. тесно) – "прижимает" молекулы АДФ и фосфат-иона, т.е. происходит синтез АТФ,
- open, O (англ. открыто) – в этом состоянии субъединица высвобождает АТФ и захватывает АДФ и ион фосфата.
При каждом обороте γ-субъединицы на 360º синтезируются три молекулы АТФ.