Репликация (синтез) ДНК происходит не беспорядочно, а в строго определенный период жизни клетки. Всего выделяют 4 фазы клеточного цикла: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap – интервал), постсинтетическую (G2).
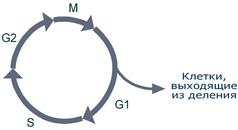
Фазы клеточного цикла
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой 35-90 кДа, уровень которых меняется в ходе клеточного цикла. По функции циклины – это активаторные субъединицы ферментов циклин-зависимых киназ (ЦЗК). Активные комплексы циклин-ЦЗК фосфорилируют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
Синтез (репликация, удвоение) ДНК происходит в S-фазу клеточного цикла, когда клетка готовится к делению. Механизм репликации, как установили Мэтью Мезельсон и Франклин Сталь в 1957 г, полуконсервативный, т.е. на каждой нити материнской ДНК синтезируется дочерняя копия.
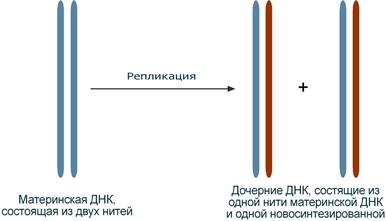
Схема полуконсервативного способа синтеза ДНК
Как любой матричный биосинтез, репликация требует наличия нескольких компонентов:
- матрица – в ее роли выступает материнская нить ДНК,
- растущая цепь – дочерняя нить ДНК,
- субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ,
- источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ,
- ферменты.
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin – начало). На каждой ДНК млекопитающих точек ori насчитывается до 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием репликативных "пузырей". В каждом таком "пузыре" имеются две репликативные "вилки", в которых происходит расплетание, раскручивание и непосредственный синтез ДНК. При этом репликативные вилки удаляются друг от друга. В целом репликация всех ДНК у эукариот заканчивается за 9 часов.
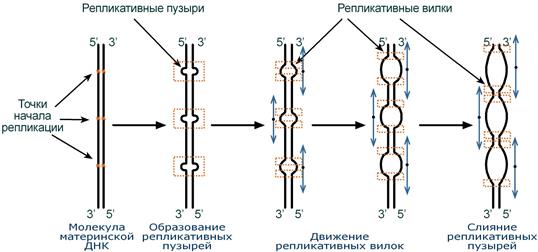
Схема возникновения репликативных пузырей и движения репликативных вилок
(нити дочерней ДНК не показаны). Стрелки - направление движения репликативных вилок.
Синтез новой цепи ДНК идет в направлении от 5'-конца к 3'-концу, т.е. 5'-конец новой ДНК остается свободным, каждый следующий нуклеотид своей 5'-гидроксильной группой присоединяется к 3'-гидроксильной группе предыдущего нуклеотида со скоростью порядка 100 штук в секунду.
В репликативной вилке в направлении 5'→3' непрерывно (т.е. обычным заурядным присоединением последующих нуклеотидов к предыдущим через С3 и С5) синтезируется только одна нить, а именно та, для которой направление синтеза совпадает с направлением движения репликативной вилки и соответствует направлению материнской нити 3'→5'. По мере расплетания ДНК и движения репликативной вилки на этой материнской нити открываются участки, где возможно безостановочное удлинение ведущей дочерней нити.
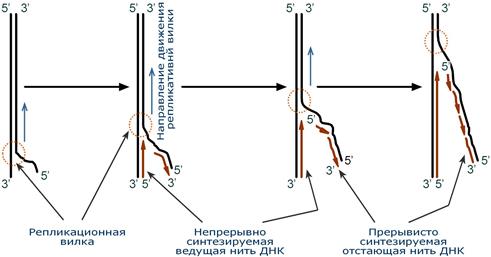
Схема движения репликативной вилки и синтеза ведущей и отстающей нитей ДНК
Направление 5'→3' для другой дочерней нити ДНК противоположно движению репликативной вилки. Поэтому синтез этой отстающей нити (в направлении 5'→3') возможен только после расплетания части ДНК и освобождения участка для синтеза.
Таким образом, синтез дочерней ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.