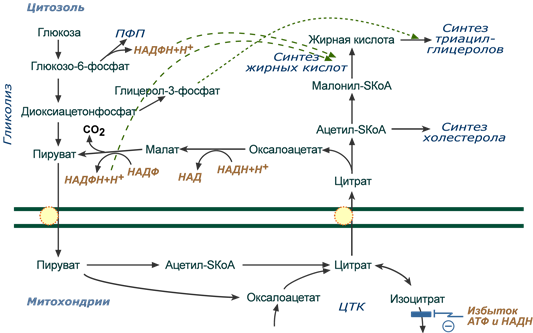
Реакции биосинтеза липидов могут идти в гладкой эндоплазматической сети клеток всех органов. Субстратом для синтеза жиров de novo является глюкоза.
Как известно, попадая в клетку, глюкоза превращается в гликоген, пентозы и окисляется до пировиноградной кислоты. При высоком поступлении глюкоза используется для синтеза гликогена, но этот вариант ограничивается объемом клетки. Поэтому глюкоза "проваливается" в гликолиз и превращается в пируват либо напрямую, либо через пентозофосфатный шунт. Во втором случае образуется НАДФН, который понадобится впоследствии для синтеза жирных кислот.
Пируват переходит в митохондрии, декарбоксилируется в ацетил-SКоА и вступает в ЦТК. Однако в состоянии покоя, при отдыхе, при наличии избыточного количества энергии в клетке реакции ЦТК (в частности, изоцитратдегидрогеназная реакция) блокируются избытком АТФ и НАДН.
Общая схема биосинтеза триацилглицеролов и холестерола из глюкозы
В результате накапливается первый метаболит ЦТК – цитрат. По градиенту концентрации он перемещается в цитозоль, расщепляется с образованием ацетил-SКоА, который далее используется в биосинтезе холестерола и жирных кислот.
Оксалоацетат, также образуемый из цитрата, восстанавливается малатдегидрогеназой до яблочной кислоты и возвращается в митохондрии
- посредством малат-аспартатного челночного механизма (на рисунке не показан),
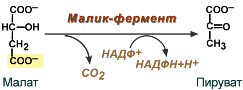
- после декарбоксилирования малата до пирувата НАДФ-зависимым малик-ферментом. Образованный НАДФН будет использован при синтезе жирных кислот или холестерина.