Сокращения и аббревиатуры, присутствующие на странице:
- p85, p110 (protein 85, protein 110) – название произошло от молекулярной массы белков 85 кДа и 110 кДа, субъединицы фермента фосфоинозитол-3-киназы,
- IRS (insulin receptor substrate) – субстрат инсулинового рецептора,
- PDK1 (3-phosphoinositide dependent protein kinase-1) – 3-фосфоинозитол-зависимая протеинкиназа-1,
- DNA-PK (DNA-dependent protein kinase) – ДНК-зависимая протеинкиназа,
- AKT1 (RAC-alpha serine/threonine-protein kinase) – серин-треониновая киназа, иначе называется протеинкиназа B,
- PIP3 (phosphatidylinositol (3,4,5)-trisphosphate) – фосфатидилинозитол-(3,4,5)-трифосфат (ФИФ3),
- Shc (Src (homology 2 domain containing) transforming protein 1) – белок семейства src-киназ, является субстратом инсулинового рецептора,
- Grb (growth factor receptor bound protein) – белок, связывающийся с рецептором фактора роста, является субстратом инсулинового рецептора,
- Ras – белок семейства ГТФ-аз, относится к онкогенам,
- GAP (GTPase activating factor) – белок, активирующий ГТФ-азную активность белка Ras,
- GEF (GTF exchange factor) – белок, ускоряющий диссоциацию комплекса Ras-ГДФ,
- SOS (son of sevenless) – белок, участвующий в обмене ГДФ на ГТФ,
- Raf -–белок, относящийся к семейству серин-треониновых протеинкиназ,
- MAPK (mitogen-activated protein kinase) – митоген-активируемая протеинкиназа,
- ERK (extracellular signal-regulated kinase) – киназа, регулируемая внеклеточным сигналом (синоним MAPK),
- MAPK-K (mitogen-activated protein kinase kinase) – киназа митоген-активируемой киназы,
- MEK (MAPK/ERK kinase) – киназа ферментов MAPK и ERK (синоним MAPK-К),
- CREB (cAMP response element-binding protein) – белок, связывающийся с цАМФ-чувствительным элементом (на ДНК),
- CRE (cAMP-response elements) – участки ДНК, регулируемые за счет изменения количества цАМФ в клетке (при участии белка CREB)/
Активация инсулинового рецептора
Рецептор инсулина представляет собой гликопротеин, построенный из двух димеров, каждый из которых состоит из α- и β-субъединиц,(αβ)2. Обе субъединицы кодируются одним геном 19 хромосомы и формируются в результате частичного протеолиза единого предшественника. Период полужизни рецептора составляет 7-12 часов. При присоединении инсулина изменяется конформация субъединиц и они связываются друг с другом, образуя микроагрегаты.
Связывание инсулина с рецептором инициирует ферментативный каскад реакций фосфорилирования. Первым делом аутофосфорилируются тирозиновые остатки на внутриклеточном домене самого рецептора. Это активирует рецептор и ведет к фосфорилированию остатков серина на особом белке, называемом субстрат инсулинового рецептора (IRS или СИР). Таких IRS выделяют четыре типа – IRS‑1, IRS‑2, IRS‑3, IRS‑4. Также к субстратам инсулинового рецептора относят белки Grb-1 и Shc, которые отличаются от IRS аминокислотной последовательностью.
Дальнейшие события могут развиваться по двум направлениям:
1. Процессы, связанные с активацией фосфатидилинозитол-3-киназы – в основном контролируют метаболические реакции обмена белков, углеводов и липидов (быстрые и очень быстрые эффекты инсулина). Сюда же относятся процессы, которые регулируют активность глюкозных транспортеров (ГлюТ) и поглощение глюкозы.
2. Реакции, связанные с активностью ферментов MAP-киназ – в целом управляют активностью хроматина (медленные и очень медленные эффекты инсулина).
Тем не менее, в клетке присутствуют ферменты, чувствительные к активации и того и другого каскадного пути.
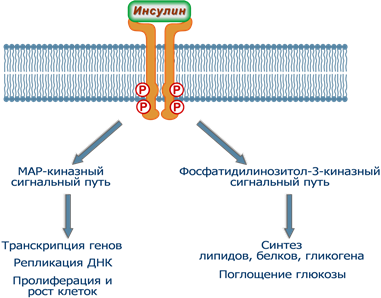
Два пути реализации эффектов инсулина
Реакции, связанные с активностью фосфатидилинозитол-3-киназы
После активации IRS-белок и целый ряд вспомогательных белков способствуют закреплению на мембране гетеродимерного фермента фосфатидилинозитол-3-киназы, содержащего регуляторную p85 и каталитическую p110 субъединицы.
Эта киназа фосфорилирует мембранный фосфатидилинозитол-4,5-дифосфат по 3-му положению до фосфатидилинозитол-3,4,5-трифосфата (PIP3). Считается, что PIP3 может выступать в качестве мембранного якоря для других элементов при действии инсулина.

Действие фосфатидилинозитолдифосфат-3-киназы на фосфатидилинозитол-4,5-дифосфат
После образования фосфатидилинозитол-3,4,5-трифосфата (PIP3) происходит активация протеинкиназы PDK-1, которая вместе с ДНК-протеинкиназой (DNA-PK) дважды фосфорилирует протеинкиназу B (AKT1), которая прикрепляется к мембране также благодаря PIP3.
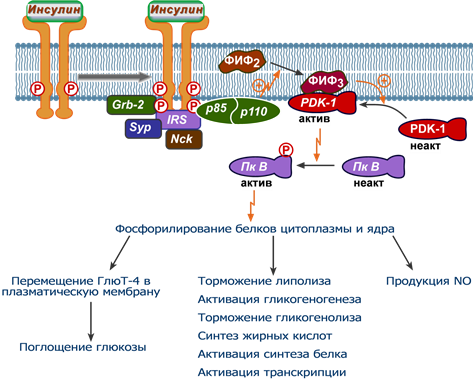
Фосфатидилинозитол-3-киназный механизм действия инсулина
Фосфорилирование активирует эту протеинкиназу В (АКТ1), она покидает мембрану и перемещается в цитоплазму и ядро клетки, где фосфорилирует многочисленные белки-мишени (более 100 штук), которые обеспечивают дальнейший клеточный ответ:
- в частности, именно действие протеинкиназы В (АКТ1) приводит к перемещению глюкозных транспортеров ГлюТ-4 на клеточную мембрану и к поглощению глюкозы миоцитами и адипоцитами,
- также, например, активная протеинкиназа В (АКТ1) фосфорилирует и активирует фосфодиэстеразу (ФДЭ), гидролизующую цАМФ до АМФ, в результате чего концентрация цАМФ в клетках-мишенях снижается. Поскольку при участии цАМФ активируется протеинкиназа А, стимулирующая гормон-чувствительную липазу и фосфорилазу гликогена, то в результате действия инсулина в адипоцитах происходит подавление липолиза, а в печени – остановка гликогенолиза.
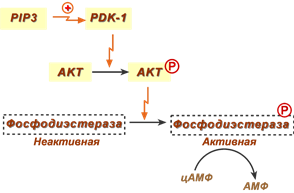
Реакции активации фосфодиэстеразы
- еще одним примером является действие протеинкиназы В (AKT) на киназу гликогенсинтазы. Фосфорилирование этой киназы инактивирует ее и в результате она не в состоянии действовать на гликогенсинтазу, фосфорилировать и инактивировать ее. Таким образом, влияние инсулина приводит к удержанию гликогенсинтазы в активной форме и к синтезу гликогена.
Реакции, связанные с активацией MAP-киназного пути
В самом начале развертывания этого пути в действие вступает еще один субстрат инсулинового рецептора – белок Shc, связывающийся с активированным (аутофосфорилированным) инсулиновым рецептором. Далее Shc-белок взаимодействует с Grb-белком и вынуждает его присоединиться к рецептору.
Также в мембране постоянно присутствует белок Ras, который в спокойном состоянии связан с ГДФ. Поблизости от Ras-белка находятся «вспомогательные» белки – GEF, SOS и белок GAP.
Формирование комплекса белков Shc-Grb активирует группу GEF-SOS-GAP и приводит к замене ГДФ на ГТФ в составе Ras-белка, что вызывает его активацию. Активный комплекс Ras-ГТФ передает активирующий сигнал на протеинкиназу Raf-1.
При активации протеинкиназы Raf-1 происходит ее присоединение к плазматической мембране, фосфорилирование дополнительными киназами по остаткам тирозина, серина и треонина, а также одновременное взаимодействие с рецептором инсулина.
Далее активированная Raf-1 фосфорилирует и активирует киназу MAPK-K (MEK), которая в свою очередь фосфорилирует следующую киназу МАPК (ERK).
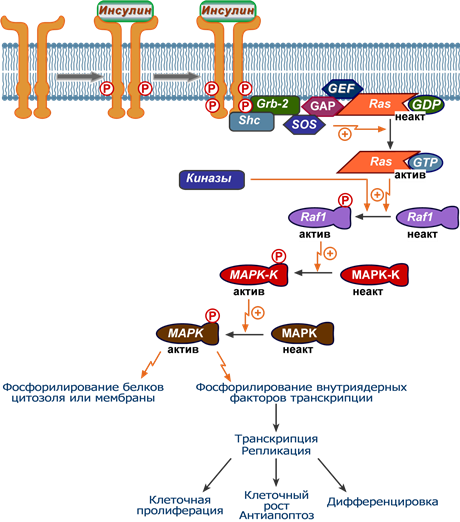
MAP-зависимый путь реализации эффектов инсулина
1. После активации МАP-киназа (МАРК) самостоятельно или при участии дополнительных киназ запускает фосфорилирование белков цитоплазмы, что изменяет их активность, например:
- активация фосфолипазы А2 приводит отщеплению от фосфолипидов арахидоновой кислоты, которая далее превращается в эйкозаноиды,
- активация рибосомальной киназы запускает процесс трансляции белков,
- активация протеинфосфатаз приводит к дефосфорилированию многих ферментов.
2. Весьма масштабной по последствиям является передача инсулинового сигнала в ядро. МАP-киназа самостоятельно фосфорилирует и этим активирует ряд факторов транскрипции, обеспечивая считывание определенных генов, важных для деления, дифференцировки и других клеточных ответов.
Одним из белков, связанных с этим механизмом, является транскрипционный фактор CREB (cAMP-response element-binding protein). В неактивном состоянии фактор дефосфорилирован и не влияет на транскрипцию. При действии активирующих сигналов фактор связывается с определенными CRE-последовательностями ДНК, усиливая или ослабляя считывание информации с ДНК и ее реализацию. Кроме MAP-киназного пути фактор чувствителен к сигнальным путям, связанным с протеинкиназой А и кальций-кальмодулином.