По сравнению с углеводами для целей накопления энергии жиры имеют преимущество - они могут накапливаться в большом количестве, не связывая воду, и в расчете на единицу веса в них содержится в два раза больше энергии. При окислении 1 грамма ТАГ образуется 9,3 ккал (38,9 кДж) тепла.
Адипоциты метаболически очень активны, быстро отвечают на гормональные сигналы. Как и другие типы клеток, адипоциты активно осуществляют гликолиз, используют цикл лимонной кислоты для окисления пирувата и жирных кислот и проводят окислительное фосфорилирование до конца. При голодании, мышечной работе, некоторых патологиях в адипоцитах активируется мобилизация жиров, когда триацилглицеролы расщепляются с образованием жирных кислот, распределяющихся в основном между печенью, скелетными мышцами и миокардом.
Запасание жира - липогенез
Запасы триацилглицеринов в адипоцитах пополняются за счет использования жирных кислот , поступающих из кишечного тракта после приема жирной пищи (переносятся в составе ТАГ хиломикронами) или жирных кислот, синтезированных из глюкозы в печени (переносятся ЛПОНП в составе ТАГ). Отрыв жирных кислот от ТАГ в липопротеинах крови обеспечивает липопротеинлипаза, находящаяся на эндотелии кровеносных капилляров. Адипоциты включают полученные жирные кислоты в состав триацилглицеринов и хранят в виде больших жировых капель (Подробно о хиломикронах и ЛПОНП).
При высоком потреблении углеводов жировая ткань также поглощает глюкозу, превращает их (через пируват и ацетил-СоА) в насыщенные и моненасыщенные жирные кислоты, Но в целом считается, что у человека синтез жиров из глюкозы происходит большей частью в гепатоцитах, а запасы в адипоцитах пополняются за счет уже готовых жирных кислот.
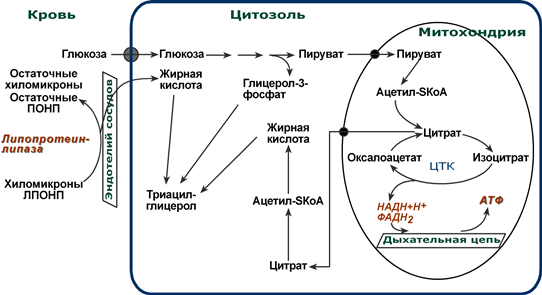
Схема метаболизма глюкозы и липидов в адипоците
Объем жирового депо у взрослого человека с нормальной массой достаточен для удовлетворения потребностей в энергии на протяжении 35-40 суток полного голодания.