При использовании углеводов, как впрочем и других веществ, перед организмом стоит две задачи – всасывание из кишечника в кровь и транспорт из крови в клетки тканей. В любом случае необходимо преодолевать мембрану.
Транспорт моносахаров через мембраны
Всасывание в кишечнике
После переваривания крахмала и гликогена, после расщепления дисахаридов в полости кишечника накапливается глюкоза и другие моносахариды, которые должны попасть в кровь. Для этого им необходимо преодолеть, как минимум, апикальную мембрану энтероцита и его базальную мембрану.
Вторично-активный транспорт
По механизму вторичного активного транспорта из просвета кишечника происходит всасывание глюкозы и галактозы. Такой механизм означает, что затрата энергии при переносе сахаров происходит, но тратится она не непосредственно на транспорт молекулы, а на создание градиента концентрации другого вещества. В случае моносахаридов таким веществом является ион натрия.
Только наличие активного транспорта позволяет перенести из внешней среды внутрь клеток практически всю глюкозу.
Фермент Na+,К+-АТФаза постоянно, в обмен на калий, выкачивает ионы натрия из клетки, именно этот транспорт требует затрат энергии. В просвете кишечника содержание натрия относительно высоко и он связывается со специфическим мембранным белком, имеющим два центра связывания: один для натрия, другой для моносахарида. Примечательно то, что моносахарид связывается с белком только после того, как с ним свяжется натрий. Белок-транспортер свободно мигрирует в толще мембраны. При контакте белка с цитоплазмой натрий быстро отделяется от него по градиенту концентрации и сразу отделяется моносахарид. Результатом является накопление моносахарида в клетке, а ионы натрия выкачиваются Na+,К+-АТФазой.
Выход глюкозы из клетки в межклеточное пространство и далее кровь происходит благодаря облегченной диффузии.
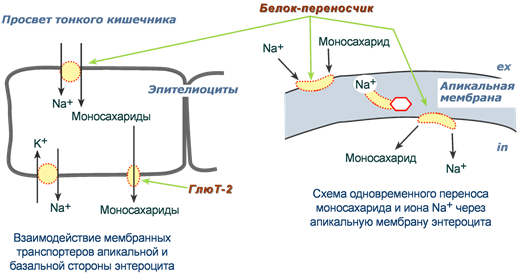
Вторично-активный транспорт глюкозы и галактозы через мембраны энтероцитов
Пассивный транспорт
В отличие от глюкозы и галактозы, фруктоза и другие моносахара всегда транспортируются белками-транспортерами, не зависящими от градиента натрия, т.е. облегченной диффузией. Так, на апикальной мембране энтероцитов находится транспортный белок ГлюТ-5, через который фруктоза диффундирует в клетку.
Для глюкозы вторично-активный транспорт используется при ее низких концентрациях в кишечнике. Если концентрация глюкозы в просвете кишечника велика, то она также может транспортироваться в клетку путем облегченной диффузии при участии белка ГлюТ-5.
Скорость всасывания моносахаридов из просвета кишечника в эпителиоцит не одинакова. Так, если скорость всасывания глюкозы принять за 100%, то относительная скорость переноса галактозы составит 110%, фруктозы – 43%, маннозы – 19%.
Транспорт из крови через мембраны клеток
После выхода в кровь, оттекающую от кишечника, моносахариды движутся по сосудам воротной системы в печень, частично задерживаются в ней, частично выходят в большой круг кровообращения. Следующей их задачей стоит проникновение в клетки органов.
Из крови внутрь клеток глюкоза попадает при помощи облегченной диффузии по градиенту концентрации с участием белков-переносчиков (глюкозных транспортеров – "ГлюТ"). Всего выделяют 12 типов транспортеров глюкозы, отличающихся локализацией, сродством к глюкозе и способностью к регулированию.
Глюкозные транспортеры ГлюТ-1 имеются на мембранах всех клеток и ответственны за базовый транспорт глюкозы в клетки, требуемый для поддержания жизнеспособности.
Особенностями ГлюТ-2 является способность пропускать глюкозу в двух направлениях и низкое сродство к глюкозе. Переносчик представлен, в первую очередь, в гепатоцитах, которые после еды захватывают глюкозу, а в постабсорбтивный период и при голодании поставляют ее в кровь. Также присутствует этот транспортер на базолатеральной мембране эпителия кишечника и почечных канальцев. Присутствуя на мембранах β-клеток островков Лангерганса, ГлюТ-2 переносит глюкозу внутрь при увеличении ее концентрации свыше 5,5 ммоль/л и выпускает из клетки наружу при снижении. Благодаря этому осуществляется тонкая регуляция сигнала для увеличения секреции инсулина.
Глют-3 обладает высоким сродством к глюкозе и представлен в нервной ткани. Поэтому нейроны способны поглощать глюкозу даже при низких ее концентрациях в крови.
В скелетных мышцах, миокарде и жировой ткани находится ГлюТ-4, только эти транспортеры являются чувствительными к влиянию инсулина. При действии инсулина на клетку они выходят на поверхность мембраны и переносят глюкозу внутрь. Указанные ткани получили название инсулинзависимых.
Некоторые ткани совершенно нечувствительны к действию инсулина, их называют инсулиннезависимыми. К ним относятся нервная ткань, стекловидное тело, хрусталик, сетчатка, клубочковые клетки почек, эндотелиоциты, семенники и эритроциты.
ГлюТ-5, как уже было сказано, находится на апикальной мембране энтероцитов, также представлен в зрелых сперматозоидах и в яичках. В первую очередь он отвечает за облегченную диффузию фруктозы.