Судьба жирной кислоты зависит от ее строения (насыщенная ЖК или полиненасыщенная ЖК) и от внутриклеточных условий (наличие или отсутствие энергии в клетке).
Абсорбтивный период
В течение нескольких часов после приема пищи (абсорбтивный период) экзогенныенасыщенные и мононенасыщенные жирные кислоты поступают с помощью хиломикронов от кишечника в ткани, которые имеют липопротеинлипазу на эндотелии капилляров (подробнее).
Параллельно в этот период времени в печени жирные кислоты способны синтезироваться из избытка экзогенной глюкозы. Далее эти эндогенные жирные кислоты этерифицируются с глицеролом в реакциях липогенеза с образованием ТАГ. Потом они транспортируются из печени в ткани, имеющие липопротеинлипазу, в составе ЛПОНП.
В клетках жировой ткани после приема пищи насыщенная жирная кислота либо синтезируется из глюкозы, либо поступает сюда из хиломикронов и ЛПОНП. Далее она направляется в липогенез и запасается в составе ТАГ.
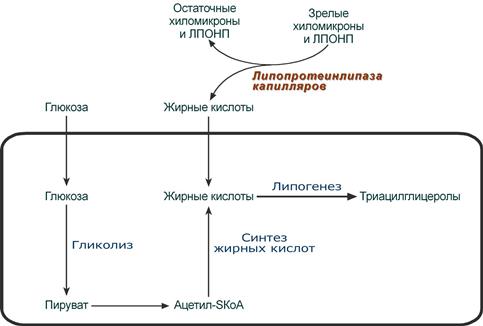
Возможные источники и пути использования жирной кислоты в абсорбтивный период
(на примере адипоцита)
Если липопротеинлипазы нет в ткани, то жирная кислота доставляется сюда в комплексе с альбумином. Этот комплекс образуется после работы липопротеинлипазы в других тканях.
Из плазмы крови внутрь клеток насыщенные жирные кислоты переносятся при участи транспортных "белков, связывающих жирные кислоты" (fatty acid binding protein, FABP). Эти белки составляют большое семейство для переноса липофильных веществ, включая эйкозаноиды и ретиноиды. Также некоторые члены семейства транспортируют липофильные молекулы от внешней клеточной мембраны к их внутриклеточным рецепторам.
Полиненасыщенные жирные кислоты (ПНЖК) обычно проникают в клетки в составе эфиров холестерола и фосфолипидов в составе ЛПВП и ЛПНП (подробнее). Эти жирные кислоты необходимы для синтеза эйкозаноидов в некоторых видах клеток, либо они участвуют в синтезе фосфолипидов для клеточных мембран.
Голодание и мышечная работа
При голодании хиломикроны и ЛПОНП в крови отсутствуют. Так как данному состоянию обычно сопутствует гипогликемия, то для ее компенсации из поджелудочной железы секретируется глюкагон. Под влиянием глюкагона и других гормонов в жировых депо активируется расщепление ТАГ до жирных кислот и глицерола (липолиз). Транспорт жирных кислот, вышедших в кровь, осуществляется альбумином.
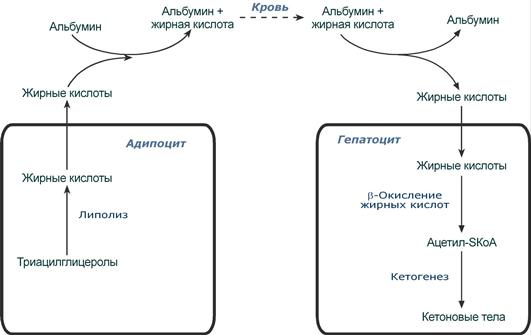
Источник и путь использования жирной кислоты при длительном голодании
(на примере гепатоцита и адипоцита)
При физической работе гормоны адреналин, соматотропин и глюкокортикоиды вызывают в адипоцитах липолиз и выход жирных кислот в кровь. В комплексе с альбумином эти кислоты доставляются, главным образом, в мышцу для обеспечения мышечного сокращения.
В состоянии покоя, когда процессы пищеварения уже закончились, при краткосрочном и длительном голодании, при физической нагрузке в большинстве клеток, кроме нейронов и эритроцитов, жирные кислоты сгорают в процессах β-окисления и ЦТК, обеспечивая 50% и более всей энергии клетки.
В печени при длительном голодании (более 20 часов) жирные кислоты направляются в кетогенез на синтез кетоновых тел. Кетоновые тела далее разносятся по организму, преобразуются в клетках в ацетил-SКоА и используются для их основного энергетического обеспечения.