Специфичность, т.е. высокая избирательность действия ферментов, основана на комплементарности структуры субстрата и активного центра фермента.
1. Стереоспецифичность – катализ только одного из стереоизомеров, например:
- специфичность к L- или D-аминокислотам – например, почти все ферменты человека взаимодействуют с L-аминокислотами,
- специфичность к цис- и транс-изомерам. Например, аспартаза реагирует только с транс-изомером – фумаровой кислотой, но не с малеиновой кислотой (цис-изомер).
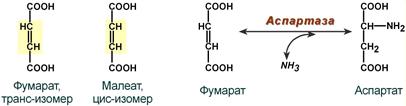
Стереоспецифичность аспартазы к транс-изомеру субстрата
2. Абсолютная специфичность – фермент производит катализ только одного вещества. Например, каталаза разрушает перекись водорода, аргиназа расщепляет только аргинин, уреаза расщепляет только мочевину, глюкокиназа фосфорилирует только D-глюкозу.
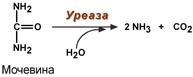
Реакция расщепления мочевины
3. Групповая специфичность – катализ субстратов с общими структурными особенностями, т.е. при наличии определенной связи или химической группы, например:
- наличие пептидной связи: • бактериальный фермент субтилизин специфичен к пептидной связи независимо от строения образующих ее аминокислот, • пепсин катализирует разрыв пептидной связи, образованной аминогруппами ароматических аминокислот, • тромбин в своих субстратах расщепляет пептидную связь только между аргинином и глицином,
- наличие α1,4-гликозидных связей в крахмале и гликогене - их гидролизует α-амилаза слюнной и поджелудочной желез,
- наличие ОН-группы: алкогольдегидрогеназа окисляет до альдегидов одноатомные спирты (этанол, метанол, пропанол).
4. Относительная групповая специфичность – превращение субстратов с некоторыми общими признаками. Например, цитохром Р450 окисляет только гидрофобные вещества, которых насчитывается около 7000.
Механизмы специфичности
В общем виде все сводится к комплементарному взаимодействию фермента и субстрата. При этом функциональные группы субстрата взаимодействуют с соответствующими им функциональными группами фермента. Наличие субстратной специфичности объясняют две гипотезы:
1. Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго соответствует конфигурации субстрата и не изменяется при его присоединении. Эта модель хорошо объясняет абсолютную специфичность, но не групповую.

Схематичное представление теории Фишера
2. Теория Кошланда (модель "индуцированного соответствия", "рука-перчатка") – подразумевает гибкость активного центра. Присоединение субстрата к якорному участку фермента вызывает изменение конфигурации каталитического центра таким образом, чтобы его форма соответствовала форме субстрата.

Схематичное представление теории Кошланда